Jika sel bekerja dalam kondisi anaerobik, ia menghasilkan energi dengan mengubah glukosa menjadi laktat dan, melalui siklus Cori, membuang yang terakhir; jika ada oksigen yang tersedia (oleh karena itu dalam kondisi istirahat), lebih dari 90% glukosa dikonsumsi secara aerobik dan hanya 10% sisanya, secara anaerobik. Ketika ada kebutuhan akan lebih banyak ATP daripada yang dapat disediakan oleh jalur aerobik (misalnya ketika otot berada di bawah tekanan), maka pasokan tambahan disediakan oleh metabolisme anaerobik (kita berada dalam kondisi kelangkaan oksigen: sesak napas, kelelahan dalam pernapasan, dll.): perlu untuk mempercepat metabolisme ini dengan mengubah laktat (yang diperoleh dari glikolisis) menjadi glukosa melalui glukoneogenesis.
Metabolisme aerobik berkembang di mitokondria.
Enzim pertama yang ditemui dalam metabolisme aerobik adalah piruvat dehidrogenase; Lebih akurat untuk mengatakan bahwa piruvat dehidrogenase adalah kompleks enzim daripada enzim, karena merupakan agregat dari 48-60 unit protein dengan tiga situs katalitik yang bekerja secara berurutan.
Piruvat dehidrogenase mengkatalisis reaksi berikut (redoks):
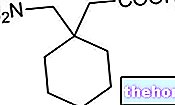
Piruvat + NAD + + CoA-SH → Asetil KoA + NADH + H + + CO2
CoA-SH adalah koenzim A: ini adalah turunan dari asam pantotenat; asetil koenzim A adalah tioester. Ini adalah proses redoks karena karbon pertama piruvat berpindah dari bilangan oksidasi tiga ke bilangan oksidasi empat (teroksidasi) dan karbon piruvat kedua beralih dari bilangan oksidasi dua ke bilangan oksidasi tiga (teroksidasi). Kemudian piruvat dioksidasi (kehilangan dua elektron sama sekali) dan NAD+ direduksi.
Seperti disebutkan, piruvat dehidrogenase memiliki tiga jenis aktivitas enzimatik, masing-masing didukung oleh kofaktor katalitiknya sendiri:
- tiamin pirofosfat (ini adalah turunan dari vitamin B1); itu aktif dalam bentuk terdeprotonasi: sebuah karbanion terbentuk.
- lipoamida (ini adalah turunan dari asam lipoat); mengandung jembatan disulfida yang sangat reaktif.
- flavin adenin dinukleotida (ini adalah turunan dari vitamin B2); itu adalah nukleotida dengan sifat redoks: pusat redoksnya terdiri dari flavin.
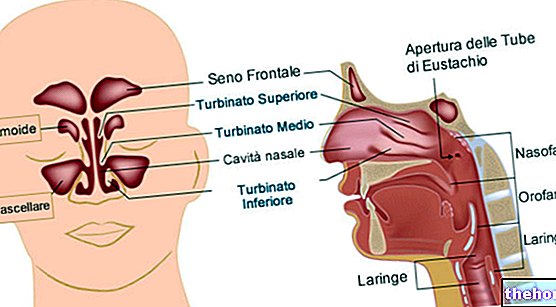
Dalam sel eukariotik, metabolisme aerobik terjadi di organel khusus sel yang merupakan mitokondria; pada bakteri metabolisme glukosa dan spesies lain terjadi di dalam sel tetapi tidak ada organel khusus.
Ketika piruvat memasuki mitokondria, ia dikenai "aksi karboksilase piruvat jika ada kebutuhan untuk melakukan glukoneogenesis (untuk merekonstruksi bahan awal), atau dapat dikenai piruvat dehidrogenase jika diperlukan untuk menghasilkan energi: " asetil koenzim A yang dibentuk oleh metabolisme aerobik merangsang aksi piruvat karboksilase, oleh karena itu, mempromosikan gluokoneogenesis dan mengurangi aksi piruvat dehidrogenase.
Sekarang mari kita lihat bagaimana piruvat dehidrogenase bekerja; pertama-tama, ada dekarboksilasi piruvat oleh aksi tiamin pirofosfat.
Lingkungan asam dapat menghambat metabolisme aerobik karena bentuk anionik tiamin pirofosfat aktif yang akan terprotonasi pada pH asam dan dekarboksilasi tidak akan terjadi.
Dekarboksilasi adalah reaksi yang sulit karena ikatan karbon-karbon harus diputus; dalam hal ini reaksi secara termodinamika disukai oleh fakta bahwa zat antara reaksi (hidroksietil-tiamin pirofosfat) memberikan resonansi (elektron p dari molekul terdelokalisasi): pirofosfat hidroksietil-tiamin ada dalam tiga kemungkinan bentuk (resonansi) dan ini membuatnya cukup stabil. Selanjutnya, hidroksietil-tiamin pirofosfat dalam bentuk anionik bertahan cukup lama untuk dapat berinteraksi dengan jembatan disulfida lipoamida (kofaktor katalitik kedua piruvat dehidrogenase); jembatan disulfida adalah lengan berosilasi (terletak di ujung rantai fleksibel yang panjang) dan dapat berpindah dari satu situs katalitik ke situs lain dalam kompleks enzim.
Kemudian lipoamida, melalui jembatan disulfida, mengikat pirofosfat hidroksietil-tiamin: asetil lipoamida diperoleh.Yang baru saja dijelaskan adalah langkah pertama dari reaksi transasetilasi yang dikatalisis oleh enzim pertama dari kompleks piruvat dehidrogenase; pada fase ini terjadi pemutusan ikatan antara gugus hidroksil dan tiamin pirofosfat yang kembali ke bentuk semula: terjadi reaksi redoks di mana jembatan disulfida bertindak sebagai oksidan (dua atom belerang tereduksi) dalam perbandingan gugus hidroksil yang teroksidasi menjadi asetil.
Setelah fase ini, lengan berosilasi dari lipoamida bergerak dan mendekati enzim kedua piruvat dehidrogenase yang melakukan aktivitas transasetilase sejati dengan membawa gugus asetil bersamanya: fase kedua dari reaksi transasetilasi yang dikatalisis oleh enzim kedua berlangsung; dengan cara ini kita telah memperoleh asetil koenzim A. Sekarang perlu untuk mengembalikan lipoamida yang dalam bentuk tereduksi: enzim ketiga piruvat dehidrogenase mengintervensi yang meredoks lipoamida dan mentransfer elektrodanya ke FAD yang direduksi menjadi FADH2. FAD / FADH2 dapat berfungsi sebagai pasangan redoks dalam dua tahap monoelektronik yang berbeda atau dalam satu tahap bielektronik tunggal.
FADH2 segera memberikan elektronnya ke NAD + memperoleh FAD dan NADH + H +.
Asetil koenzim A, diperoleh seperti yang dijelaskan, adalah produk awal untuk siklus Krebs (atau siklus asam trikarboksilat).