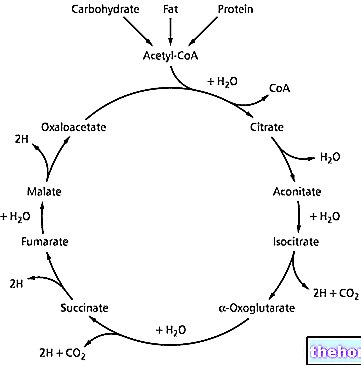
Glikogen adalah makromolekul (massa molekul sekitar 400 juta dalton) dari -glukosa di mana terutama ada ikatan -1,4 glikosidik dan percabangan dalam rasio 1:10, karena ikatan -1,6 glikosidik.
Glikogen merupakan bahan cadangan dan terus terdegradasi dan dilarutkan; di seluruh massa sel tubuh, ada sekitar 100 g glikogen: sebagian besar berada di hati di mana ia dapat bergerak dan, oleh karena itu, dapat digunakan sebagai cadangan untuk organ lain (glikogen dalam otot tidak bergerak).
Enzim-enzim yang mengkatalisis degradasi dan sintesis glikogen semuanya ada di dalam sitoplasma, oleh karena itu diperlukan sistem regulasi yang membuat satu jalur menjadi tidak aktif ketika jalur yang lain aktif: jika ada glukosa, jalur tersebut diubah menjadi glikogen (anabolisme) yang cadangan, sebaliknya, jika c "diperlukan untuk glukosa, maka glikogen terdegradasi (katabolisme).

Enzim yang terutama terlibat dalam pemecahan glikogen adalah glikogen fosforilase; enzim ini mampu membelah ikatan -1,4 glikosidik menggunakan ortofosfat anorganik sebagai agen litik: pembelahan terjadi dengan cara fosforolitik dan diperoleh glukosa 1-fosfat.
Pada lima atau enam unit dari titik percabangan, enzim glikogen fosforilase tidak lagi dapat bekerja sehingga terlepas dari glikogen dan digantikan oleh enzim pengurai yang merupakan transferase: di situs katalitik enzim ini c "adalah" histidin yang memungkinkan transfer tiga unit sakarida ke rantai glikosidik terdekat (histidin menyerang karbon pertama dari molekul glukosa). Enzim yang baru saja disebutkan adalah glikosiltransferase; Pada akhir kerja enzim ini, hanya satu unit glukosa yang tersisa pada rantai samping dengan karbon pertama terikat pada karbon keenam dari glukosa pada rantai utama Unit glukosa terakhir pada rantai samping dilepaskan oleh kerja enzim ini. "enzim -1,6 glikosidase (enzim ini merupakan bagian kedua dari enzim deramifikasi); mengingat bahwa cabang-cabang dalam glikogen dalam rasio 1:10, dari degradasi lengkap makromolekul kita memperoleh sekitar 90% glukosa 1-fosfat dan sekitar 10% dari glukosa.
Kerja enzim-enzim yang disebutkan di atas memungkinkan penghapusan rantai samping dari molekul glikogen; aktivitas enzim-enzim ini dapat diulang sampai degradasi lengkap rantai terjadi.
Mari kita pertimbangkan hepatosit; glukosa (diasimilasi melalui makanan), ketika memasuki sel diubah menjadi glukosa 6-fosfat dan dengan demikian diaktifkan. Glukosa 6-fosfat, dengan aksi fosfoglukomutase, diubah menjadi glukosa 1-fosfat: yang terakhir adalah prekursor biosintesis non-langsung; dalam biosintesis, bentuk gula yang diaktifkan digunakan yang diwakili oleh gula yang terkait dengan difosfat: biasanya uridildifosfat (UDP). kemudian diubah menjadi UDP-glukosa, metabolit ini di bawah aksi glikogen sintase yang mampu mengikat UDP-glukosa ke ujung non-pereduksi dari glikogen yang sedang tumbuh: diperoleh glikogen memanjang dari unit glukosidik dan UDP.UDP diubah oleh enzim difosfokinase nukleosida menjadi UTP yang kembali ke sirkulasi.
Degradasi glikogen terjadi oleh aksi glikogen fosforilase yang melepaskan molekul glukosa dan mengubahnya menjadi glukosa 1-fosfat. Selanjutnya, fosfoglukomutase mengubah glukosa 1-fosfat menjadi glukosa 6-fosfat.
Glikogen disintesis, terutama di hati dan otot: dalam organisme terdapat 1-1,2 hektogram glikogen yang didistribusikan ke seluruh massa otot.
Glikogen miosit merupakan cadangan energi hanya untuk sel ini sedangkan glikogen yang terkandung dalam hati juga merupakan cadangan untuk jaringan lain, yaitu dapat dikirim, sebagai glukosa, ke sel lain.
Glukosa 6-fosfat yang diperoleh di otot dari degradasi glikogen kemudian dikirim, dalam hal kebutuhan energi, ke glikolisis; di hati, glukosa 6-fosfat diubah menjadi glukosa oleh aksi glukosa 6-fosfat fosfatase (karakteristik enzim hepatosit) dan dibawa ke dalam aliran darah.
Glikogen sintase dan glikogen fosforilase keduanya bekerja pada unit non-pereduksi glikogen sehingga harus ada sinyal hormonal yang memerintahkan aktivasi satu jalur dan memblokir yang lain (atau sebaliknya).
Di laboratorium dimungkinkan untuk memperpanjang rantai glikogen dengan memanfaatkan glikogen fosforilase dan menggunakan glukosa 1-fosfat dalam konsentrasi yang sangat tinggi.
Dalam sel, glikogen fosforilase hanya mengkatalisis reaksi degradasi karena konsentrasi metabolit sedemikian rupa sehingga menggeser kesetimbangan reaksi berikut ke kanan (yaitu menuju degradasi glikogen):

Mari kita lihat mekanisme kerja glikogen fosforilase: oksigen asetal (yang bertindak sebagai jembatan antara unit glukosa) mengikat hidrogen dari fosforil: reaksi antara terbentuk diberikan oleh karbokation (pada glukosa yang semuanya " ekstremitas) yang mengikat fosforil (Pi) dengan sangat cepat.
Glikogen fosforilase membutuhkan kofaktor yaitu piridoksal fosfat (molekul ini juga merupakan kofaktor untuk transaminase): ia memiliki fosforil terprotonasi sebagian (piridoksal fosfat dikelilingi oleh lingkungan hidrofobik yang membenarkan keberadaan proton yang terikat padanya) . Fosforil (Pi) mampu mentransfer proton ke glikogen karena fosforil ini kemudian memperoleh kembali proton dari fosforil piridoksal fosfat yang terprotonasi sebagian. Probabilitas bahwa, pada pH fisiologis, fosforil kehilangan protonnya dan tetap terdeprotonasi sepenuhnya sangat rendah.
Sekarang mari kita lihat bagaimana fosfoglukomutase bekerja.Enzim ini menyajikan, di situs katalitik, residu serin terfosforilasi; serin menghasilkan fosforil menjadi glukosa 1-fosfat (di posisi enam): glukosa 1,6-bifosfat terbentuk untuk waktu yang singkat, kemudian serin difosforilasi ulang dengan mengambil fosforil di posisi satu. Phosphogluco mutase dapat bekerja dua arah, yaitu mengubah glukosa 1-fosfat menjadi glukosa 6-fosfat atau sebaliknya; jika glukosa 6-fosfat diproduksi, dapat dikirim langsung ke glikolisis, di otot, atau diubah menjadi glukosa di hati.
Enzim uridyl phosphogluco transferase (atau UDP glukosa pyrophosphorylase) mengkatalisis reaksi transfer glukosa 1-fosfat ke UTP melalui perlekatan pada fosforil a.
Enzim yang baru saja dijelaskan adalah pirofosforilasi: nama ini disebabkan oleh fakta bahwa reaksi yang berlawanan dengan yang baru saja dijelaskan adalah pirofosforilasi.
Glukosa UDP, diperoleh seperti yang dijelaskan, mampu memperpanjang rantai glikogen, dengan unit monosakarida.
Hal ini dimungkinkan untuk membuat reaksi berkembang menuju pembentukan glukosa UDP dengan menghilangkan produk yang pirofosfat; enzim pirofosfatase mengubah pirofosfat menjadi dua molekul ortofosfat (hidrolisis anhidrida) dan dengan demikian, menjaga konsentrasi pirofosfat tetap rendah sehingga membuat proses pembentukan glukosa UDP secara termodinamika disukai.
Seperti disebutkan, glukosa UDP, berkat aksi glikogen sintase, mampu memperpanjang rantai glikogen.
Konsekuensinya (dalam perbandingan 1:10) disebabkan oleh fakta bahwa, ketika rantai glikogen terdiri dari 20-25 unit, enzim bercabang (memiliki "histidin pada situs katalitiknya) campur tangan, mampu mentransfer serangkaian 7-8 unit glikosidik lebih jauh ke hilir dari 5-6 unit: dengan demikian percabangan baru dihasilkan.
Untuk alasan asal gugup atau jika energi diperlukan karena aktivitas fisik, adrenalin dikeluarkan dari kelenjar adrenal.
Sel target adrenalin (dan noradrenalin) adalah hati, otot, dan jaringan adiposa (pada yang terakhir terjadi degradasi trigliserida dan sirkulasi asam lemak: akibatnya, glukosa diproduksi di mitokondria 6-fosfat, menjadi dikirim ke glikolisis, sedangkan di adiposit, glukosa 6-fosfat diubah menjadi glukosa oleh aksi enzim glukosa 6-fosfatase dan diekspor ke jaringan).
Mari kita lihat, sekarang modalitas aksi adrenalin. Adrenalin berikatan dengan reseptor yang ditempatkan pada membran sel (miosit dan hepatosit) dan ini menentukan translasi sinyal dari luar ke dalam sel. Protein kinase diaktifkan yang bekerja secara simultan pada sistem yang mengatur sintesis dan degradasi glikogen:
Glikogen sintase ada dalam dua bentuk: bentuk terdefosforilasi (aktif) dan bentuk terfosforilasi (tidak aktif); protein kinase memfosforilasi glikogen sintase dan memblokir aksinya.
Glikogen fosforilase dapat ada dalam dua bentuk: bentuk aktif di mana serin terfosforilasi hadir dan bentuk tidak aktif di mana serin didefosforilasi. Glikogen fosforilase dapat diaktifkan oleh enzim glikogen fosforilase kinase. Glikogen fosforilase kinase aktif jika terfosforilasi dan tidak aktif jika terdefosforilasi; protein kinase memiliki substrat glikogen fosforilase kinase, yaitu, ia mampu memfosforilasi (dan, oleh karena itu, mengaktifkan) yang terakhir yang, pada gilirannya, mengaktifkan fosforilase glikogen.
Setelah sinyal adrenalin berakhir, efeknya pada sel juga harus berakhir: enzim fosfatase kemudian mengintervensi spesies protein.


.jpg)